Comparative Metabolomic Analysis Reveals Stress-Resilient Metabolites in Dwarf Tomato Plant
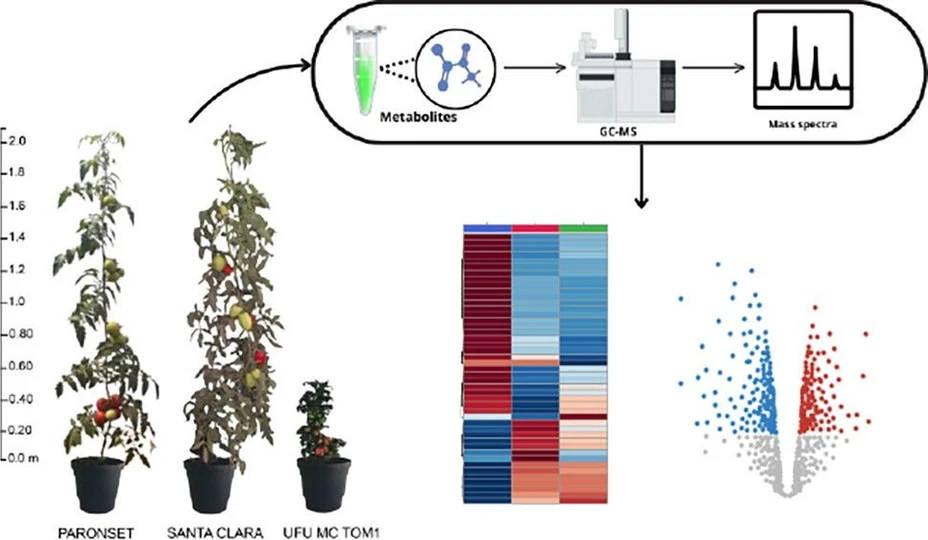
ACS Agric. Sci. Technol. 2026, 6, 1, 83–91: Graphical abstract
This study compares the metabolomic profiles of dwarf and commercial tomato cultivars using nontargeted GC–MS metabolomics. The dwarf cultivar UFU TOM MC 1 showed elevated levels of osmoprotectants, antioxidants, and stress-responsive amino acids, including myo-inositol and proline.
Pathway analysis revealed enrichment in carbohydrate, amino acid, and protein metabolism pathways associated with stress adaptation. These findings highlight the potential of dwarf tomato cultivars for breeding programs focused on improving stress tolerance, resilience, and nutritional quality.
The original article
Comparative Metabolomic Analysis Reveals Stress-Resilient Metabolites in Dwarf Tomato Plant
Brena R. M. Ikehara*, Natália R. Almeida, Gabriel M. Maciel, Ana C. S. Siquieroli, Camila S. de Oliveira, Lucas M. Pereira, Ana L. A. Ribeiro, and Frederico G. Pinto
ACS Agric. Sci. Technol. 2026, 6, 1, 83–91
https://doi.org/10.1021/acsagscitech.5c00253
licensed under CC-BY 4.0
Selected sections from the article follow. Formats and hyperlinks were adapted from the original.
Tomato (Solanum lycopersicum) is one of the most important vegetable crops globally, extensively cultivated and consumed across diverse regions. In 2022, global production was estimated at about 180 million tons, of which roughly 39 million tons were processed by the food industry. (1) Belonging to the Solanaceae family, along with other important vegetables, tomato fruits offer several health benefits due to their rich range of phytonutrients, including carotenoids, flavonoids, and phenolic acids, and also contribute to strengthening the agroeconomic chain. (2,3) This fruit is considered a model species due to their diploid genome, filling a niche in the studies of fleshy fruits that cannot be occupied by Arabidopsis. (4)
Tomato plants can be cultivated in temperate to hot climatic zones, but significant amounts of agricultural input and irrigation are required. However, they can be harshly affected by biotic and abiotic stresses such as drought, salinity, extreme temperatures, and attacks by herbivores or pathogens, compromising productivity and fruit quality. (5) The impacts of global warming on temperatures threaten the development and production of many crops worldwide, including tomato plants, and therefore food and nutrition security. (6) For example, low humidity triggers calcium depletion and increases soil salinity, leading to plant diseases and greatly reducing the tomato crop yield. High temperatures also affect important steps for fruit development, including fruit setting, flowering, and ripening, further reducing tomato yield. (7,8)
To reduce the consequences of these changes on tomato production, several breeding programs use wild species, such as S. pimpinellifolium, S. chileno, S. peruvianum, S. pennellii, and S. hirsutum, along with improved varieties S. lycopersicum var. Lycopersicum, for crosses that resulted in enhanced biotic and abiotic resistances. In the search for resistance genes in tomatoes, plants with dwarfism genes have stood out in recent years. (9,10) Besides being easy to work with, due to their monogenic recessive heredity, they are known for being capable of bringing positive morphological and physiological changes, such as smaller plant architecture, shorter internodes, resistance to pests (11,12) and diseases, (13) and saline and water stress. (14)
While recognizing the benefits of tomato plants with the dwarfism gene, such as the production of hybrids with higher yield and resistance, (15) there are still gaps in the germplasm of this species regarding their chemical composition, including primary and secondary metabolites in leaflets, fruits, and roots. Addressing these gaps requires research focusing on elucidating the chemical composition of tomato varieties, which allows describing their nutritional potential. In order to achieve this, metabolomics, a set of analytical techniques applied to identify substantial amounts of metabolites in biological samples, is employed. Specifically, a nontargeted metabolomics approach, through analyses in gas chromatography coupled to mass spectrometry (GC-MS), has the potential to help better understand the plant metabolome and explain the correlation of metabolites with quality and yield, generating a greater understanding of the physiological and biochemical processes of the dwarf tomato plant. (16,17)
This research aimed to investigate the chemical composition of leaflets from different tomato cultivars through GC-MS metabolomic analysis. The main goal was to better understand the differences and advantages of genetic variation, adaptability, and resilience of dwarf and commercial tomato cultivars through metabolomic analysis of their leaflets.
Experimental Section
Gas Chromatography-Mass Spectrometry Analyses
The analysis was performed using gas chromatography coupled with a mass spectrometer (GCMS-QP2010, Shimadzu, Kyoto, Japan), equipped with a DB-5MS capillary column (30 m × 250 μm internal diameter). The sample injection temperature used was 280 °C, with a constant helium gas flow of 1.0 mL/min throughout the process. The initial temperature for chromatographic separation was 80 °C, which was maintained for 2 min. Subsequently, the temperature was increased by 5 °C every minute, up to 290 °C, which was maintained for 5 min. The injection volume was 1 μL with a split of 20:1. The interface and ion source temperatures used were 250 °C. The detector voltage was 1.2 kV, and the electron impact ionization mode was selected for the ionization of metabolites at 70 eV. An alkane standard (C10–C40) was used for the quality control and retention index calculation.
Results
The metabolomic analysis on GC-MS generated the chromatogram (Figure S1), in which 45 compounds with significantly differentially expressed among UFU MC TOM 1, Santa Clara, and Paron Set (Figure 1A) were annotated, according to analysis of variance (ANOVA) (Fisher’s LSD test, p < 0.05, Table S1) and shown as a heatmap (Figure 1B). Notably, the metabolomic profile of the UFU MC TOM 1 cultivar is statistically different from the others, as confirmed by Partial Least Squares Discriminant Analysis (PLS-DA) with values of R2 = 0.9940 and Q2 = 0.7928 (Figure 1C). PLS-DA analysis, using compounds with significant differences in their abundances, showed a distinct clustering for each group, with components 1 and 2 explaining 91.4 and 5.4% of the total variance, respectively (Figure S2).
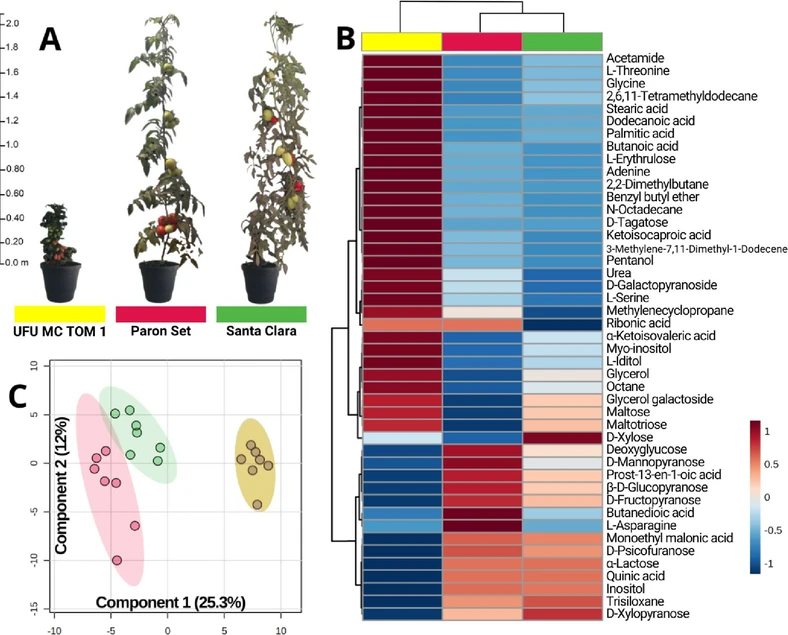 ACS Agric. Sci. Technol. 2026, 6, 1, 83–91: Figure 1. (A) Visual comparison of the three cultivars. (B) Heatmap of the mean intensity levels across the analyzed groups: UFU MC TOM 1 (yellow), Paron Set (red), and Santa Clara (green). (C) PLS-DA plot, accounting for 37.3% of the observed variation in group separation.
ACS Agric. Sci. Technol. 2026, 6, 1, 83–91: Figure 1. (A) Visual comparison of the three cultivars. (B) Heatmap of the mean intensity levels across the analyzed groups: UFU MC TOM 1 (yellow), Paron Set (red), and Santa Clara (green). (C) PLS-DA plot, accounting for 37.3% of the observed variation in group separation.
The divergent compounds between the groups underwent pathway analysis enrichment, based on the Kyoto Encyclopedia of Genes and Genomes (KEGG) database. The ten metabolic pathways identified as significantly impacted were galactose metabolism (p = 0.0003); valine, leucine, and isoleucine biosynthesis (p = 0.0023); cyanoamino acid metabolism (p = 0.0051); glyoxylate and dicarboxylate metabolism (p = 0.0051); glycine, serine, and threonine metabolism (p = 0.0074); sulfur metabolism (p = 0.0145); butanoate metabolism (p = 0.0185); alanine, aspartate, and glutamate metabolism (p = 0.0302); and biosynthesis of unsaturated fatty acids (Figure 2).
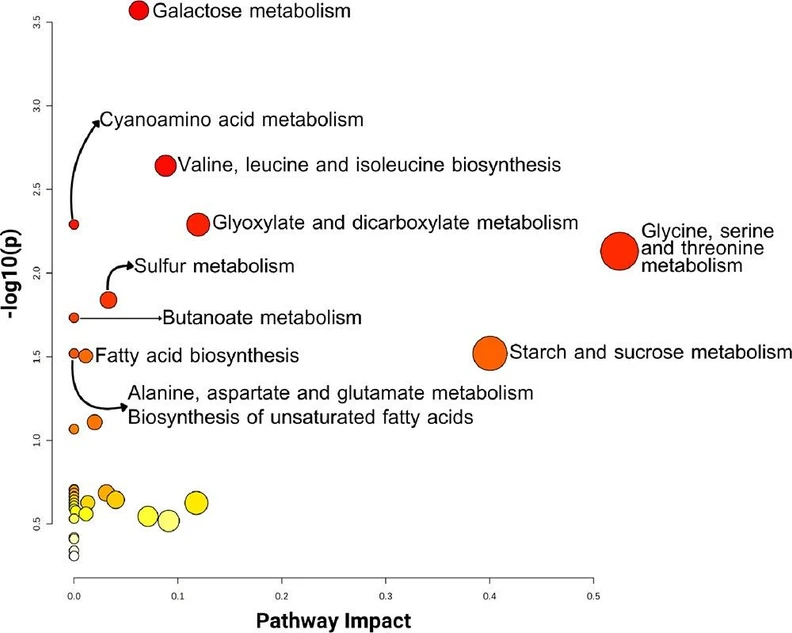 ACS Agric. Sci. Technol. 2026, 6, 1, 83–91: Figure 2. Metabolic pathway analysis overview. The analysis took into consideration the significant metabolites identified previously in this work. The color of the circles represents −log(p), with red being the highest, while the size of each circle is proportional to the impact of the pathway.
ACS Agric. Sci. Technol. 2026, 6, 1, 83–91: Figure 2. Metabolic pathway analysis overview. The analysis took into consideration the significant metabolites identified previously in this work. The color of the circles represents −log(p), with red being the highest, while the size of each circle is proportional to the impact of the pathway.
Discussion
Several metabolites were significantly more abundant in UFU MC TOM 1, revealing distinct metabolic profiles among the evaluated cultivars. These metabolites encompass various chemical classes, primarily amino acids, carbohydrates, and organic acids. As the experiment was carried out under controlled and uniform conditions, without the imposition of abiotic or biotic stress, the observed variations represent relative quantitative distinctions between cultivars. Consequently, potential links to stress responses or other biological functions are inferred from the literature rather than demonstrated experimentally.
Previous studies have demonstrated that the UFU TOM MC 1 cultivar displays broad-spectrum resistance to a variety of pests, with these defense responses being largely attributed to elevated acyl sugar concentrations in the leaf tissue. (11,12) The metabolite profile identified in UFU TOM MC 1 indicates the presence of metabolic pathways potentially linked to resistance against multiple biotic stresses, highlighting its value for future functional and breeding studies. Currently, tomato pest management remains largely dependent on insecticide application. (53) The insights gained from this study indicate that advancing the development of pest-resistant tomato cultivars could markedly decrease the reliance on pesticides, promoting more sustainable production systems. Previous studies have reported UFU MC TOM 1 as resistant to Xanthomonas euvesicatoria pv perforans, but the underlying mechanisms remain unclear. (13)
The pronounced differences in metabolites and metabolic pathways between UFU TOM MC 1 and commercial cultivars offer important evidence of the superior resilience to biotic and abiotic stresses of this dwarf tomato lineage. In general, the accumulation of osmoprotectants, antioxidants, and stress-responsive amino acids, along with the activation of defense-related pathways, sheds further light on the dwarf genetic resource for breeding programs aimed at improving crop resilience under changing environmental conditions.
Although untargeted metabolomics does not replace the rigorous validation protocols typical of targeted quantitative assays, it provides a robust and widely accepted approach for detecting meaningful metabolic differences. This strategy enables the identification of variations that are more likely to reflect genuine biological differences. While some metabolites may have nutritional relevance, the experimental design does not allow definitive conclusions on this parameter. Although the findings do not provide direct evidence of resistance mechanisms, they advance our understanding of the biochemical and physiological bases of the resilience of UFU TOM MC 1 and generate testable hypotheses about its adaptive potential, which may support tomato productivity, especially in the context of global climate change.