ATR-FTIR Assessment of the Antioxidant and Anti-Inflammatory Effects of (−)-Epicatechin in MASLD
ACS Omega 2026, 11, 23, 33333–33344: Graphical abstract
This study investigates the antioxidant and anti-inflammatory effects of (−)-epicatechin in an in vitro model of metabolic dysfunction-associated steatotic liver disease (MASLD). Lipid peroxidation, inflammatory, and profibrotic markers were assessed alongside ATR-FTIR spectroscopy to evaluate whether spectral changes reflect disease progression and treatment response.
Epicatechin reduced lipid peroxidation products, TNF-α, and TGF-β1 in a concentration-dependent manner, with stronger protective effects observed during pretreatment. ATR-FTIR spectral changes closely correlated with biochemical markers, demonstrating its potential as a rapid, non-destructive tool for monitoring oxidative stress, inflammation, and fibrosis in MASLD research.
The original article
ATR-FTIR Assessment of the Antioxidant and Anti-Inflammatory Effects of (−)-Epicatechin in MASLD
Marija Hefer, Nikolina Filipović, Stjepan Šarić, Nela Malatesti, Ana Petrović, Elvira Kovač-Andrić*, and Martina Smolić*
ACS Omega 2026, 11, 23, 33333–33344
https://doi.org/10.1021/acsomega.5c09978
licensed under CC-BY 4.0
Selected sections from the article follow. Formats and hyperlinks were adapted from the original.
Metabolic dysfunction-associated steatotic liver disease (MASLD) represents one of the most prevalent chronic liver disorders. Due to its predisposition to progress toward metabolic dysfunction-associated steatohepatitis (MASH), fibrosis, cirrhosis, and even hepatocellular carcinoma (HCC), MASLD has become a significant global health burden and a major challenge for healthcare systems. (1−4)
The majority of patients with MASLD or MASH are obese and frequently present with additional metabolic comorbidities, most commonly insulin resistance, type 2 diabetes mellitus, and dyslipidemia. (5−7) Furthermore, most conditions associated with MASLD and MASH are characterized by lipid metabolism impairment, with elevated levels of oxidative stress. Impaired lipid metabolism, together with prolonged oxidative stress, promotes hepatocellular lipotoxicity and mitochondrial dysfunction. These alterations affect the antioxidant defense system and increase reactive oxygen species (ROS) generation, thereby contributing to the onset of MASLD and its progression to MASH, along with more advanced liver diseases. (8,9)
One of the main results of oxidative stress is lipid peroxidation, in which ROS attack (poly)unsaturated fatty acids within membrane lipids. This initiates a series of structural and functional alterations, where the formation of conjugated dienes (CDs) reflects early double-bond rearrangements in fatty acyl chains, while lipid hydroperoxides (LOOHs) represent key propagation-phase products that destabilize cell membranes and increase oxidative injury. (10,11) Therefore, determining LOOH and CD levels, along with their overall LOOH/CD ratio, might be useful for determining the lipid peroxidation dynamics and the progression of MASLD.
Along with oxidative damage to the cell structures, inflammation is also a key regulator of MASLD progression. In this context, tumor necrosis factor-α (TNF-α) is suspected to play a central role as its levels are found to increase alongside MASLD to MASH progression. (12−14) It has also been found that ROS and TNF-α are closely interconnected, where elevated TNF-α levels could serve as an indicator of oxidative cellular damage and enhanced lipid peroxidation. (15−17) Often, one of the results of MASLD progression is the development of liver fibrosis, which is usually mediated by transforming growth factor beta 1 (TGF-β1) as its levels are frequently elevated in MASH and contribute to the worsening of hepatic steatosis. (18) Figure 1 summarizes the pathways associated with MASLD to MASH progression, with emphasis on lipid peroxidation (CDs, LOOHs) and inflammatory/fibrogenic signaling (TNF-α, TGF-β1).
 ACS Omega 2026, 11, 23, 33333–33344: Figure 1. Schematic representation of lipid peroxidation and its role in MASLD progression. Lipid accumulation in MASLD leads to the release of free radicals and ROS, which attack (poly)unsaturated fatty acids within membrane lipids, leading to lipid peroxidation, inflammation (via TNF-α), and fibrotic changes (via TGF-β1), thereby contributing to the transition from MASLD to MASH and further progression to cirrhosis and HCC. In lipid peroxidation, reactive radicals (R·) subtract the hydrogen from the (poly)unsaturated lipid, thereby resulting in the formation of a lipid radical, which undergoes rearrangements to form a CD. The lipid radical then reacts with molecular oxygen to form an unstable lipid peroxyl radical, which further subtracts a hydrogen from another (poly)unsaturated lipid, thereby resulting in the formation of a LOOH. The subtraction of hydrogen from the (poly)unsaturated lipid also results in the formation of a lipid radical, thereby perpetuating the chain reaction. The figure was created with Servier Medical Art (https://smart.servier.com/) and MarvinSketch (ChemAxon, https://chemaxon.com/marvin).
ACS Omega 2026, 11, 23, 33333–33344: Figure 1. Schematic representation of lipid peroxidation and its role in MASLD progression. Lipid accumulation in MASLD leads to the release of free radicals and ROS, which attack (poly)unsaturated fatty acids within membrane lipids, leading to lipid peroxidation, inflammation (via TNF-α), and fibrotic changes (via TGF-β1), thereby contributing to the transition from MASLD to MASH and further progression to cirrhosis and HCC. In lipid peroxidation, reactive radicals (R·) subtract the hydrogen from the (poly)unsaturated lipid, thereby resulting in the formation of a lipid radical, which undergoes rearrangements to form a CD. The lipid radical then reacts with molecular oxygen to form an unstable lipid peroxyl radical, which further subtracts a hydrogen from another (poly)unsaturated lipid, thereby resulting in the formation of a LOOH. The subtraction of hydrogen from the (poly)unsaturated lipid also results in the formation of a lipid radical, thereby perpetuating the chain reaction. The figure was created with Servier Medical Art (https://smart.servier.com/) and MarvinSketch (ChemAxon, https://chemaxon.com/marvin).
However, the exact role of different lipid peroxidation indicators and their contribution to MASLD progression toward MASH have not yet been fully elucidated, with the number of studies addressing this issue remaining limited. Given these limitations in research, attention has increasingly turned toward dietary antioxidants, particularly polyphenols or mixtures of polyphenol-rich plant extracts, as potential modulators of oxidative stress and lipid peroxidation in hepatic steatosis. (19−23)
Among available polyphenols, we selected (−)-epicatechin (EC) because of its strong antioxidant and radical scavenging capacity, with its documented effects on inflammatory signaling. (24−26) One study even found that the antifibrotic properties of EC were comparable to those of silymarin, with EC showing even stronger reducing effects on malondialdehyde (MDA) formation in vivo. (27) Since MDA is a secondary product of LOOH breakdown, (28) these results could link EC’s potential antifibrotic properties to its ability to attenuate lipid peroxidation, thereby potentially mitigating the progression of MASLD to MASH. Unlike more extensively studied compounds such as epigallocatechin gallate, (29,30) EC has been less investigated in the context of MASLD, providing an opportunity to explore a novel candidate. At the same time, the growing use of advanced spectroscopic techniques offers new opportunities to monitor biochemical alterations in cells and tissues.
Attenuated total reflection-Fourier transform infrared (ATR-FTIR) spectroscopy recently emerged as a promising tool for monitoring such changes. The application of ATR-FTIR spectroscopy and similar techniques for steatosis and lipid quantification, along with determining the lipid peroxidation extent, has been introduced, especially in the context of hepatic steatosis. Determining the intensity of C═O vibrational stretching at ∼1740 cm–1 (triacylglycerols, cholesteryl esters, lipid oxides), as this type of bond usually occurs in lipid peroxidation, could also be valuable for determining the overall extent of steatosis. (31−33) Along with lipid peroxidation extent, determining the degree of unsaturation in MASLD models is crucial for understanding the changes in cellular components as the degree of unsaturation is often found to decrease due to steatotic changes in cells and tissues. (Poly)unsaturated fatty acids contribute to membrane fluidity and are essential for maintaining membrane integrity and signaling, while their depletion leads to increased membrane rigidity and impaired cellular function. (34,35) Given their high susceptibility to peroxidation, a reduction in their levels could indicate either an adaptive response to oxidative stress or a disruption of lipid homeostasis. Therefore, correlating the degree of unsaturation (∼3010 cm–1 for ═C–H) and the intensity of carbonyl stretching (∼1740 cm–1 for C═O) with the LOOH/CD ratio and cytokines, TNF-α and TGF-β1, could be valuable for the noninvasive monitoring of disease progression.
Methods
Experimental Design
Two experimental models were introduced for the experiments: EC pretreatment and EC Posttreatment models. In both models, the untreated cells (UT) group was used as a negative (vehicle) control group, while the sodium oleate-treated group (OA) was used as a positive control group, with EC (control) being the highest concentration of EC used as a control for the tested compound. In the Pretreatment model, the cells were pretreated with 10, 30, and 50 μM EC for 4 h, followed by a 24 h posttreatment with 1.5 mM sodium oleate. In the Posttreatment model, the cells were treated with 1.5 mM sodium oleate for 12 h, followed by the addition of the same concentrations of EC in the presence of 0.75 mM sodium oleate. All of the experimental groups were labeled as EC50+OA, EC30+OA, and EC10+OA, according to the concentration used in the experiments. EC concentrations of 10, 30, and 50 μM were chosen based on our previous MTS assay and Oil Red O staining results, where we tested 10–100 μM EC to evaluate its effect on MASLD in the Pretreatment model. (42) As it was proven effective in the Pretreatment model, the Posttreatment model was added to evaluate whether EC could also mitigate oxidative and inflammatory/fibrotic changes once steatosis was already established. Another important consideration was to assess the potential cytotoxicity of the concentrations used, as EC and other polyphenols are often consumed as dietary supplements with poorly regulated dosages. As these cytotoxic effects are often subtle and overlooked in conventional biochemical assays, combining FTIR spectral analysis with complementary assays could provide a more comprehensive assessment of potential cytotoxicity.
ATR-FTIR Assessment of Lipid Peroxidation
ATR-FTIR spectroscopy was used to determine the extent of lipid peroxidation and steatosis. The lipid samples were dissolved in chloroform and normalized to a lipid concentration of 20 mg/mL. After the evaporation of the solvent, the ATR-FTIR spectra were recorded using an Agilent Cary 630 FTIR spectrophotometer (Agilent Technologies, Santa Clara, CA, USA), at the Faculty of Biotechnology and Drug Development at the University of Rijeka. All ATR-FTIR spectra were processed using the automatic least-squares baseline correction algorithm implemented in Agilent MicroLab Expert v.10.5 software (4000–650 cm–1; clean scans: 4; threshold: 0.002; background scans: 2; sample scans: 100; resolution: 16 cm–1; apodization: HappGenzel; phase correct: Mertz; method gain: 221). Spectra of pure chloroform were recorded under identical conditions and subtracted from sample spectra to remove any residual solvent contributions. Chloroform was recorded to be excluded in the background, and the data were collected in the spectral range 3200–1600 cm–1 on an ATR device. To ensure comparability between samples, the spectra were normalized against the asymmetric CH2 stretching vibration of lipid acyl chains (2925–2922 cm–1), which was used as an internal reference band due to its relative stability across treatments and minimal alteration during lipid peroxidation. Baseline correction was applied prior to normalization to minimize background variability. Relative intensities of lipid peroxidation-associated bands (C═O stretching at 1744–1735 cm–1 and ═C–H stretching at 3010 cm–1) were then expressed as a ratio to the CH2 reference peak, allowing quantitative comparison of the lipid peroxidation extent between experimental groups. The data were presented as percent relative intensities (%Irel).
Results and Discussion
ATR-FTIR Spectroscopy for the Assessment of Lipid Peroxidation and Steatosis Extent
To evaluate the overall steatosis and lipid peroxidation extent in HepG2 cells, ATR-FTIR spectroscopy was used. Figure 5 shows the 3200–1600 cm–1 spectral region, including the key C═O (1744–1735 cm–1) and ═C–H (3010 cm–1) stretching bands, which correlated with the biochemical parameters measured in this study. The increased intensity of the C═O band in sodium oleate-treated groups corresponds to the accumulation of oxidized lipid species derived from peroxidation of (poly)unsaturated fatty acids. On the other hand, a decrease in the ═C–H stretching band near 3010 cm–1 indicates a progressive depletion of double bonds which undergo rearrangement to form CDs or further transformations associated with lipid peroxidation.
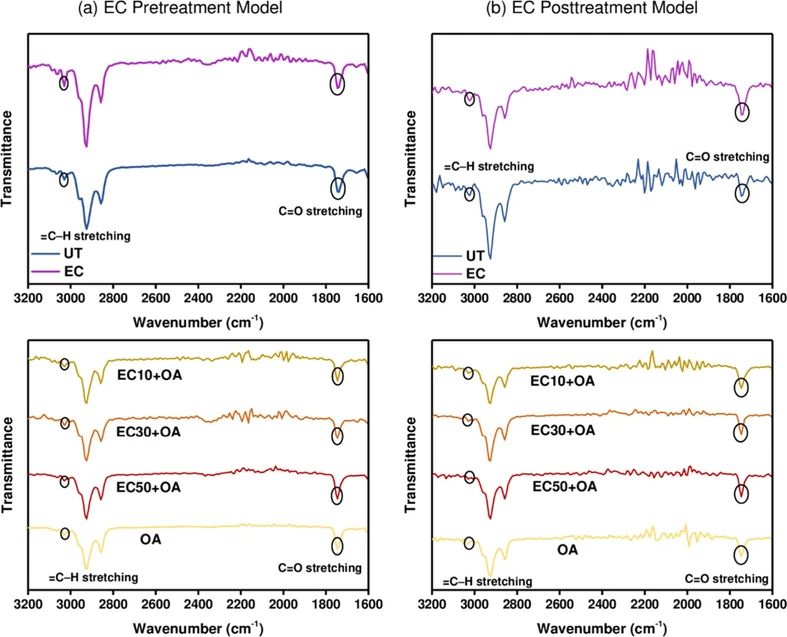 ACS Omega 2026, 11, 23, 33333–33344: Figure 5. ATR-FTIR spectra of control groups: UT (blue) and EC (control) (purple); along with experimental groups: OA (yellow), EC50+OA (red), EC30+OA (orange), and EC10+OA (brown) in the wavenumber range 3200–1600 cm–1 for the (a) EC Pretreatment Model and (b) EC Posttreatment Model.
ACS Omega 2026, 11, 23, 33333–33344: Figure 5. ATR-FTIR spectra of control groups: UT (blue) and EC (control) (purple); along with experimental groups: OA (yellow), EC50+OA (red), EC30+OA (orange), and EC10+OA (brown) in the wavenumber range 3200–1600 cm–1 for the (a) EC Pretreatment Model and (b) EC Posttreatment Model.
All experimental groups show similar FTIR profiles in the lipid regions; however, to determine the overall extent of lipid peroxidation and steatosis, the relative intensities of ═C–H stretching of unsaturated lipids (3010 cm–1) and C═O stretching of esters (1744–1735 cm–1) were measured according to the baseline and normalized according to asymmetric CH2 stretching of lipid chains (2925–2922 cm–1) (Figure 6). This approach was chosen as the greatest changes were noticed among the given intensities of the peaks in all groups.
 ACS Omega 2026, 11, 23, 33333–33344: Figure 6. Percent relative intensities (%Irel) of ═C–H stretching (unsaturated lipids) and C═O stretching of esters for the (a) EC Pretreatment Model and (b) EC Posttreatment Model. The results are presented as mean values of %Irel ± SD.
ACS Omega 2026, 11, 23, 33333–33344: Figure 6. Percent relative intensities (%Irel) of ═C–H stretching (unsaturated lipids) and C═O stretching of esters for the (a) EC Pretreatment Model and (b) EC Posttreatment Model. The results are presented as mean values of %Irel ± SD.
Also, as indicated in the literature, the increase in intensities of C═O stretching and a decrease in intensities of ═C–H stretching are mostly associated with an increase in the lipid peroxidation extent. (33,50) Furthermore, a decrease in ═C–H stretching corresponds to a decrease in the (poly)unsaturated fatty acid content, which is often noted in MASLD/MASH models. (51,52)
Conclusion
In a sodium oleate-induced hepatic steatosis model, EC showed a concentration-dependent effect on the overall radical scavenging ability of HepG2 cells. However, when EC was given as a posttreatment, these results were not achievable. EC also reduced propagation-phase lipid peroxidation by reducing relative LOOH levels in both models, lowering the LOOH/CD ratio (except for the EC50+OA pretreatment group), and effectively decreasing sodium oleate-induced increases in TNF-α and TGF-β1 concentrations. ATR-FTIR results were consistent with changes in lipid (per)oxidation and reflected the overall antioxidant and anti-inflammatory effects of EC observed through biochemical assays. Compared with biochemical assays, ATR-FTIR spectroscopy has several advantages, including rapid and nondestructive analysis that requires minimal sample preparation. Considering that it can simultaneously capture multiple molecular changes, this represents a reliable approach for accurate and complementary insights into lipid peroxidation and redox alterations in MASLD. However, it is important to correlate ATR-FTIR spectral changes with biochemical assays to further strengthen its interpretation.